


Charles M. Schweik
Over the past decade, considerable attention has been given to the depletion of species in forests of the developing world (Myers, 1988; Aldhous, 1993; Repetto, 1988; Lovejoy, 1980; Task Force on Global Biodiversity, 1989; Norton, 1986; Reid and Miller, 1989). Often these studies take a macro view of the problem, focusing on general political or economic influences (Repetto, 1988; Richards and Tucker, 1988). A second research area shifts attention to the individual and searches for deeper understanding of influential variables that drive foraging behavior. Some of this literature focuses on the influence of institutions or rules in use that create or modify foraging incentives (Ascher, 1995; McKean, 1992; Thomson, Feeny, and Oakerson, 1992; and Appendix I to this volume).
Often micro-level studies utilize the forest area as a whole as the unit of analysis in which the researcher develops some measure of forest condition. Typically, these studies will (1) take a sample of vegetation using forest plots, (2) calculate species abundance indicators30 from these plots, and (3) use these indicators to describe the current status of the forested area as a whole. If the researcher is fortunate, prior data on the forest exists and these newly generated indicators can be compared to ones developed from previous time periods. General conclusions can then be made regarding the change in forest resources over time and the impact of current institutional arrangements governing the area.
While these forest-level analyses often produce quite valuable information regarding forest condition and forest governance as a whole, some information -perhaps important information-may be lost due to the aggregation of forest plot data. A hypothetical example may drive home this point. Suppose the abundance measure for a particular important species31 of a forested area is determined to be quite high. A deeper spatial analysis32 may reveal that the majority of these species are found in locations far from forager households.33 At a forest level of analysis, the researcher concludes that the sustainability of the important species appears to be adequate. But from a plot level of analysis-a spatial perspective-an overharvesting problem exists. In this instance, aggregation yields false conclusions.
In addition, from an institutional perspective, aggregation to the forest level may lead to a lack of understanding of the foraging dynamics within a community. For example, the existing spatial distribution of a species may reflect the community's response to established harvesting rules. Designated protected areas that are effectively monitored may exhibit species abundance measures higher than in other unprotected areas within the same forest. To the researcher trying to understand how governance arrangements influence the harvesting behavior of foragers, a spatial analysis could be quite revealing.
The primary goal of this chapter is to develop a new methodologyusing recent advances in Geographic Information System (GIS) technologies and maximum likelihood regression -to identify patterns in species distribution that differ from what we would expect in a natural, undisturbed34 setting. The development of such a methodology helps analysts in two ways. First, it provides a way to identify forest depletion using data collected from only one time period. Second, it helps analysts understand the influence of institutional arrangements on species harvesting across space. The identification of spatial anomalies in a distribution help identify harvester reaction to the established institutional structure.
To achieve this goal, this study utilizes data from a recent International Forestry Resources and Institutions (IFRI) data collection effort in Nepal. Specifically, the study investigates two questions:
1. Does the spatial pattern of an important species found in a Nepali forest differ from what we would expect in a "natural," undisturbed setting?
2. Given the discovery of a disturbance in the spatial distribution of the species, what accounts for this outcome?
The chapter is organized in the following manner: First, I describe the study site, the species of interest, and the process used to collect the forest plot data. Second, I establish the theoretical foundation that guides the development of a "species density" model. This model-referred to as a trend surface regression model-contains a spatial component to address the research questions identified above. At this point, I also describe the operationalization of all influential variables. Third, I provide an overview of the statistical methods used. Specifically, I report the techniques used for identifying the natural, undisturbed spatial distribution of the species of interest and the results of the maximum likelihood regression. Fourth, I discuss findings of particular importance, and then conclude the chapter with some suggestions for future research.
In October 1994, forested areas within the Shaktikhor Village Development Committee (VDC)35 in the Chitwan District of southern Nepal were chosen for a study of forest governance. The project, a part of the larger IFRI research program, gathered information regarding forest management, use, and condition along with socioeconomic attributes of villages that utilize these resources (see Appendix I to this volume). A research team comprised of Nepali researchers36 and the author spent six weeks living in the Shaktikhor area.
Ten settlements or villages comprise the Shaktikhor community, eight of which are located along the banks of two converging rivers (see Map 5.1). The majority of the villagers are subsistence farmers and depend heavily on forest products for their livelihood. Three forests37 exist in the area: Sugabhanjyang, Latauli, and Kaswang. The villagers utilize these forests daily as sources of fodder, fuelwood, food products, and timber for construction activities.
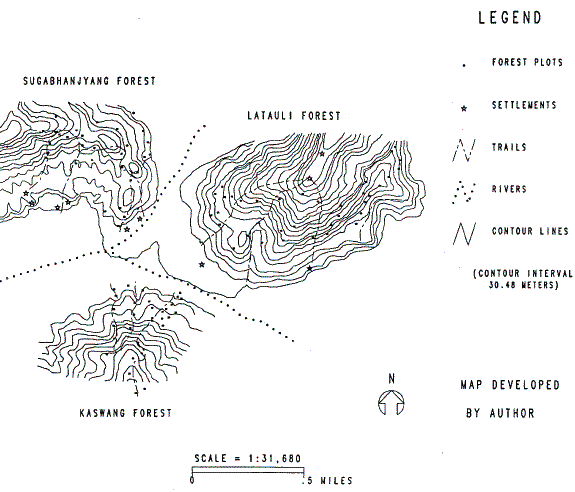
For each of these forests, the team utilized ten-meter radius circular forest plots for sampling. Due to the steep terrain within these forests, the team followed trails to reach 50-meter altitudinal intervals. At each vertical location, a random number was used to determine the direction and the distance from the trail that the corresponding plot should be taken. Overall, 97 forest plots were sampled. Data recorded for each plot included:
All three forests are under the jurisdiction of His Majesty's Government District Forest Office (DFO). However, some community developed rules appear to exist. Detailed information was collected on specific rules in use-both rules as specified by the DFO as well as a few rules established and in use by local communities. This is discussed in further detail below.
In order to investigate research question 1-the identification of unusual patterns in the distribution of an important species-this section estimates a maximum likelihood regression model. But before proceeding to a description of this statistical method, a brief theoretical discussion of important variables will be provided. In addition, for each theoretically important factor identified, the corresponding IFRI variable used to operationalize the concept will be presented.
The variable we are trying to explain in this study is the abundance of an important species within forest plots. The most important species reported by villagers (IFRI, 1994) in the Shaktikhor community is Shorea robusta, locally known as Sal. This species provides fuelwood for cooking, and timber for house and tool construction. Sal leaves are also highly valued for use in religious ceremonies. For these reasons, Shorea robusta is chosen as the species for analysis.
There are a variety of methods for measuring the abundance of Shorea robusta species in a particular plot. Species frequency, density, and importance value are measures widely used in the forestry literature and each could be an appropriate index. For reasons described more fully in the methodology section, the density (the number of individuals) of Shorea robusta trees per plot will be used as the dependent variable in the model.
Many factors determine whether a species is found in a particular area. These influential factors can be divided into three categories: (1) Plot abiotic stresses, (2) Plot biotic stresses, and (3) Human stresses.38
Abiotic stresses. Each forest plot contains physiographic characteristics that influence the capacity for particular species to grow in its environment. These characteristics include slope, aspect (or slope orientation), elevation, and soil type and condition. These attributes play a tremendous role in the number and type of species that exist within a plot (Spun and Barnes, 1992).
Slope steepness, aspect, and elevation: These are crucial factors for determining whether a species exists in a given forest plot. These variables dramatically influence the ecosystem characteristics-exposure to sunlight, rainfall, etc. -that exist within the plot. Any model attempting to identify the important factors for species growth must control for these features. The field team recorded slope steepness with a clinometer and elevation with an altimeter. Global Positioning System equipment and topographic maps helped to cross-check elevation readings.
Soil type and condition: Soil nutrients, moisture, and physical composition are also highly influential factors in the growth and survival of particular species. Three soil horizons are typically reported in soil analysis: the "o" horizon (humus or ground litter layer), the "a" horizon (a darker mineral layer at the top of the soil), and the "b" horizon (the soil deeper in the ground). The color of these layers as well as textures (sandy, loamy or clayey) are also important determinants in what species can grow in the area. A soil analysis was performed on each forest plot and included depth, color, and texture of these horizons (IFRI, 1994).
Other natural disturbances: Other naturally occurring disturbances influence what grows in a particular area. A plot area may be subject to severe weather damage caused by a lightning strike or a fallen tree. This damage results in renewed competition by the existing vegetation that survived. The IFRI forms record observations for each plot on natural disturbances, but very few disturbances were identified at the Shaktikhor site.
Blotic stresses. The number of a particular species within a plot is also influenced by a variety of biotic environmental stresses.
Species competition: Competing species in the forest plots are highly influential in determining whether another species can survive in a particular location. Competition for light, moisture, and nutrients exists in every plot, and species that can best tolerate the plot's conditions with respect to other species will have the best opportunity to grow. Since all trees in each plot were identified, measured, and entered into the IFRI database, this analysis uses a summation of the diameter at breast height (DBH) for species other than Shorea robusta within each plot.
Seed trees: Proximity of seed trees often determine if a tree will grow in a particular plot. The type of seed and its transportation medium influence where it may grow. The seed of a Shorea robusta is a samara39 and may be carried a great distance by wind disturbances (Storrs and Storrs, 1990). While this is a difficult if not impossible concept to operationalize, this influence can be partially captured using data from forest plots that are identified as "neighbors." For this reason, a count of Shorea robusta neighbor trees in the most adjacent plot was included in the model.
Animal grazing: Animal activity also influences species survival within a plot. Animals forage for particularly tasty or nutritious species and these grazing habits often then determine the fate of many seedlings. In contrast, species not particularly interesting to animals may continue to survive or even thrive. The IFRI forms capture this concept by recording any signs of grazing found in forest plots. One possible limitation of this data is that it only captures the existence of grazing activities undertaken recently. However, many wild species forage from a central location (Stephens and Krebs, 1986) and consequently fresh evidence of grazing may capture long-term foraging in the area.
Animals also play a role in how species are replicated. Certain species have seeds that attach to an animal's fur or are consumed and deposited via droppings in other locations (Spurr and Barnes, 1992). Unfortunately, the data collected do not allow for an operationalization of this phenomenon.
Human stresses. A human's decision to forage or not to forage in a particular location is another important influence on whether a species exists in that location. Every forest plot exhibits additional characteristics that determine whether humans will harvest its resources.
Ease of access to plot (plot location): Optimal foraging theory developed by anthropologists depicts human foragers as actors who maximize their net rate of return of energy per unit of foraging time (Smith, 1983). While a number of alternative theories on foraging decision making exist (see Smith, 1983: 627), they all characterize the forager as a person who strives to minimize his or her search time and effort (Hayden, 1981; Winterhalder, 1993). What optimal foraging suggests is that the location of a plot in relation to villages may be an important determinant of the existence of Shorea robusta species. If humans harvest a particular species at a rate higher than can be regenerated by the forest, optimal foraging suggests that fewer individuals of that species will be found in locations easily accessed by humans (e.g., a short distance away from the village, near a path, or at a low elevation). The model captures this influence by including plot location.
In order to determine the location of forest plots, I created a three-dimensional representation of the forested area and produced X, Y, and Z coordinates for each plot. Two technologies-Global Positioning System (GPS40) and GIS -have made the collection and storage of accurate plots coordinates much easier. To create these data, I digitized contour lines from topographic sheets of the area and created a GIS representing real-world coordinates. Of the 97 forest plots sampled, GPS recorded the location of 31. I transferred these point locations onto the real-world contour GIS. I digitized the other 66 plot locations into the GIS using field maps, plot elevations recorded on IFRI forms, and personal knowledge of the field site and the location of trails. The result of this process was a plot location GIS (the output of which is Map 5.1) that is capable of identifying accurate plot locations and distances (in meters) relative to one another. I then utilized the GIS to produce X, Y, and Z (elevation) coordinates for each plot that are used as independent variables in the regression model.
Property rights and species rules-in-use: Smith (1983) reports that empirical studies on optimal foraging theory have revealed that in some instances human foragers are selective in their utilization of available resources. Other studies have revealed foragers who exhibit much less concern. Smith also states that there is little agreement in the anthropological community over these foraging differences (1983: 628-29). While not stated specifically, Smith's discussion alludes to the importance of community relationships and the important role institutional arrangements play in the influence of human foraging patterns and their efforts for natural resource preservation.41
Ostrom (1990) extends Smith's argument by emphasizing the role of effective institutional arrangements in the use and management of natural resources. Institutions in this context refer to the working rules that govern the harvesting of a particular species or particular areas (what we might refer to as management units) within a forest. In the latter case, one particular plot may exhibit a higher density of a particular species in part due to the existence of rules that have set that particular area aside as a reserve. Foragers may bypass the plot area because these rules are accepted by the community.
Monitoring and sanctioning practices: Rules by themselves may not be particularly useful unless monitoring and sanctioning mechanisms exist that effectively enforce them (Nostrum, 1990). For example, a particular plot may be subject to better monitoring practices because of its proximity to a forest ranger post. Sanctioning differences may exist between plots due to established management units. A harvesting infraction in an area exhibiting a particularly delicate landscape (e.g.., susceptible to landslides) may result in a stiffer penalty than an infraction in an area not susceptible to such disasters.
The IFRI forms obtain information about a large number of variables that encompass these concepts (Nostrum et al., 1994). Some institutional arrangements have been established at two different levels: the DFO (national) level and the VDU (local) level.
The DFO has an interest in the area because all three forests are designated DFO-owned land. There are three formally established DFO rules related to forest product use in this region. First, anyone who is a member of a VDU is permitted to harvest grass, tree fodder, and deadwood from forests within that VDU to support their daily subsistence requirements. Second, live tree harvesting is permitted only if formal permission is received from the DFO prior to harvesting. Third, a "no encroachment" rule exists that prohibits the conversion of DFO forest land to some other land use.
These rules are enforced by DFO forest guards who patrol an area greater than 100 square kilometers. Much of this area is extremely steep and hilly. Only one road exists, running just north and adjacent to the river below the Sugahanjyang Forest. This road runs from the western border of Map 5.1 and stops at the juncture of the two rivers. This road plays an important component to the rule monitoring conducted by DFO guards. The only DFO rule enforcement we witnessed during our weeks in the field occurred along this western road. Villagers report very little interaction with the guards, particularly in areas on the eastern side of the river juncture. Overall, DFO monitoring appears to be quite infrequent and, when it does occur, it is usually in the area of the Sugabhanjyang Forest adjacent to the road.
In recent years, His Majesty's Government of Nepal has established a policy where forest areas are being transferred as communal forests owned and governed by community members (Pardo, 1993). The local VDU (Shaktikhor) is in the process of convincing the DFO to transfer ownership of these forests to these communities. To convince the DFO that their interest is genuine, the VDU has established one additional forest rule-that VDU permission must be obtained before any harvesting of live timber trees (including Shorea robusta) is undertaken. To monitor and enforce this rule, the VDU expects villagers in the community to report any violations they see. Villagers report that this rule is rarely, if ever, followed. No conflicts between villagers over rule enforcement has been reported. In addition, we witnessed fresh evidence of timber harvesting and the villagers accompanying us exhibited little concern over the infraction.
Thus, in regard to formal institutions in the area, my conclusion is that while rules do exist and are understood by villagers, they lack adequate enforcement. Live Shorea robusta are essentially open access species in all forests of the area (IFRI, 1994).
In addition to these formal rules that govern these forests, there exists one unwritten, de facto rule or what some might refer to as a social norm. Some villagers of lower social status (caste) are not permitted to harvest in the Sugabhanjyang Forest (see Map 5.1). Villagers of higher caste live in settlements closer to the western Sugabhanjyang Forest (north-west side of Map 5.1). The people living in villages adjacent to the Latauli Forest (north-east side of Map 5.1) are primarily of a lower social caste. The higher castes on the western side of the site report that they frequently harvest from both the western Sugabhanjyang and eastern Latauli Forests.
The lower caste villagers however, who reside on the eastern Latauli side, report that they only forage in the Latauli Forest and never in the Sugabhanjyang Forest. While no conflicts were reported over infringements of this unwritten rule, social pressures from high-caste villages adjacent to the Sugabhanjyang Forest appear to keep these lower caste people out of this forest. This suggests that the Latauli side may be subject to more harvesting pressure than the Sugabhanjyang Forest.
The location variables described earlier-the X, Y coordinatesoperationalize this social relationship. If the Latauli side is subject to greater harvesting due to foraging behavior in response to this understood rule, these location variables will reflect that influence. No other institutional arrangements exist in regard to forest harvesting in the Shaktikhor area.42 Figure 5.1 summarizes these theoretical influences described in this section.
Number of Individuals of a Species in a Forest Plot |
= F |
(Abiotic stresses, Biotic stresses, Human stresses) |
|
||
In this section, I apply multiple regression estimation to the model shown in Figure 5.1. Traditional Ordinary Least Squares (OLS) regression assumes the underlying distribution of the dependent variable to be a normal, bell-shaped curve. Ludwig and Reynolds (1988) state that a normal distribution assumption is often not correct when counts of biological phenomena are utilized. King (1989) argues that maximum likelihood estimation (MLE) is a better approach, for it allows the researcher to specify both the distribution of the dependent variable and the relationship independent variables have with the dependent variable (referred to as the "functional form" or the model).43
The first assumption required for MLE is the identification of the most theoretically appropriate distribution of the dependent variable. This distributional flexibility of MLE avoids residual violation problems that traditional OLS estimation encounters. The dependent variable-the number of Shorea robusta species in a plot-is an event-count variable.44 MLE requires the identification of the distribution of this event-count variable, and our investigation requires that the distribution be in a setting that is relatively undisturbed by human activities.
Counts of biological species usually follow one of three types of spatial arrangements: random, clustered, or uniform (Ludwig and Reynolds, 1988). In the case of a random dispersal of species, each plot has an equal chance of hosting a Shorea robusta individual, resulting in a frequency distribution that is normally distributed (centered around the mean). In such random patterns, the variance will be very close to the mean in value. The second pattern, a clustered pattern, is commonly found in biological studies and follows a negative binomial distribution. Ulustering will result in a large number of plots where no Shorea robusta individuals are identified. The variance in a clustering pattern will be greater than the mean. Finally, the third pattern often identified is a uniform pattern where almost every plot exhibits the same number of Shorea robusta individuals. In these spatial patterns, the variance will be less than the mean (Ludwig and Reynolds, 1988).
A "reference forest" was used to identify the natural distribution of the Shorea robusta species.45 A reference forest is a forest that (1) is an adequate representation of the other forests of interest and (2) is generally undisturbed by human activity.
Three forests were sampled in the Shaktikhor area. The Kaswang Forest (found in the southern area of Map 5.1) satisfies the above two conditions. First, it is an adequate representation of the other two forests for they all are identified as Shorea robusta climax forests. While their general slope aspects do vary slightly, they all contain significant Shorea robusta populations. Second, the Kaswang Forest appears to be the one that exhibits the least exposure to human foraging.
Four reasons, based on IFRI (1994) findings, support the above conclusion that Kaswang is subject to minimal human foraging. First, the forest's location in relation to the villages in the area minimizes its use. For much of the year, high and strong flowing waters of the Kair River cut off the Kaswang Forest from the settlements. Even in the dry season, the river can be quite difficult to cross. Second, only about half of the villages report any harvesting of any species in the Kaswang Forest. In addition, the villagers harvesting from Kaswang report that they utilize it only as a secondary source of forest products. The other two forests supply their primary needs. Third, no evidence in discussions with villagers reveals any past dramatic change (e.g., fire, mass clear-cutting, etc.) in any of the three forests. Therefore, Kaswang has not been subject to a major catastrophe. Fourth, an examination of importance values of Shorea robusta trees and saplings between the three forests (Table 5.1) reveals that overall, the dominance, density, and frequency of the Shorea robusta species is higher in Kaswang than in the other two forests. This suggests it is under much less harvesting pressure than the other two. For these reasons, I utilize the 31 forest plots sampled in the Kaswang Forest separately as the reference forest to determine a natural distribution of Shorea robusta.
The variance-to-mean ratio or index of dispersion test (Ludwig and Reynolds, 1988) identifies the natural distribution of the Shorea robusta count for the Kaswang Forest (Table 5.2). The value for the Uhi-squared statistic (df 30) is larger than the critical value at the .01 probability level, implying that Shorea robusta in natural settings follows a clumped pattern (variance is greater than the mean). This suggests that the dependent variable, the number of Shorea robusta species per plot, follows a negative binomial distribution in an undisturbed setting (Ludwig and Reynolds, 1988: 24). Accordingly, a negative binomial is the appropriate distributional assumption for maximum likelihood estimation for this analysis.
Sugabhanjyang |
Latauli |
Kaswang (Reference Forest) | |
Shorea robusta Trees |
.2725 |
.1937 |
.4912 |
Shores robusta Saplings |
.0446 |
.0589 |
.2768 |
Average Number of Individuals per Plot |
5.968 |
Number of Plots |
31 |
Variance |
12.644 |
Index of Dispersion (variance/mean ratio) |
2.119 |
X2 statistic [ X2 = ID(N-1) ] |
63.562* |
*Fp <.01
The second assumption MLE requires is the specification of the model's functional form. This requires the researcher to specify relationships (e.g., linear or nonlinear) between the dependent variable and each independent variable. An assumption of strict linearity is reasonable for specifying the relationships in Figure 5.1. There is no theoretical justification for the inclusion of exponential components. With the two assumptions for MLE regression specified, a negative binomial maximum likelihood was estimated.
Maximum Likelihood Estimation: The results of the MLE are presented in Table 5.3. Caution is required when interpreting the regression coefficients. Since they are a result of a negative binomial regression, they cannot be interpreted in the same manner as coefficients would be if they were produced by an OLS regression. One of the most intuitive ways of interpreting these results is by creating the incident rate ratio (IRR). IRRs can be easily interpreted as a percentage of growth or decline in the dependent variable due to a one-unit change in the independent variable, holding all else constant.
The steepness of the slope is found to have a slight negative influence on the existence of Shorea robusta species. Holding everything else constant, a one-degree increase in the steepness of the plot will result in approximately a three (1-IRR) percent decrease in the number of Shorea robusta trees. This makes intuitive sense. The larger the angle of the forest plot, the more difficult it would be for trees to develop a strong foundation. The slight influence of slope, however, is not surprising. Treks through these forests revealed very steep slopes exhibiting an abundance of vegetation. During the field visit, many species, Shorea robusta being one, appeared to be quite capable of establishing a foundation regardless of the steepness of the slope.
The orientation of the slope (slope aspect) is found to be weakly statistically significant. While this does suggest that Shorea robusta does have aspect preferences, the statistical weakness of the test is not unexpected. These forests are Shorea robusta dominated climax forests and the species is found in all types of terrain facing all types of directions.
Only one soil variable, the depth of the humus layer, was found to have any statistically significant explanatory relationship with the existence of Shorea robusta trees. This may reflect some problem in the soil data collected in this site. The soil analysis was conducted without a soil color chart. While the same people collected all of the soil data, the ability to discern soil color or texture accurately may have been lacking. The other possibility is that because Shorea robusta is in fact the climax species of these forests, it may be robust in terms of its ability to grow in a variety of soils. The depth of the humus layer may be significant because of its relationship to the Shorea robusta trees itself. The humus layer reflects, in part, tree leaf litter, and therefore should be correlated with the existence of trees.
The existence of competing tree biomass is negatively related to the existence of Shorea robusta trees in the plot and found to be highly statistically significant. The coefficient is, however, quite small. This reflects the measure used for competing biomass -the sum of the DBH in centimeters. A one-centimeter increase in competing tree DBH results in a less than one percent decline in the number of Shorea robusta species. This makes intuitive sense: the existence of large competing trees in a micro eco-system will produce an area not receptive to Shorea robusta growth.
Independent Variables |
Coefficients |
IRR |
Slope Steepness |
-.0287*** (.010) |
.9716 |
Slope Aspect |
.1610* (.092) |
1.175 |
Elevation |
.0043 (.001) |
1.004 |
Humus Layer Depth |
-.3559*** (.126) |
.7005 |
A and B Horizon Depth |
-.0086 (.029) |
.9914 |
A and B Horizon Color |
.059 (.068) |
1.061 |
A Horizon Texture |
-.1736 (.179) |
.8406 |
Competing Tree Biomass |
-.0044*** (.001) |
.9956 |
# of Shorea robusta Neighbors |
-.1526*** (.056) |
.8584 |
Signs of Livestock Grazing |
-.4884 (.404) |
.6136 |
X Coordinate |
-.0008** (.0002) |
.9991 |
Y Coordinate |
.0006 (.0005) |
1.0006 |
Intercept |
.8062 (1.35) |
Negative Binomial log-likelihood: - 103.61
n=66
***p < .01
**p < .05
*p < .10
The number of Shorea robusta trees in the nearest-neighbor plot was also found to be influential at the .01 level. At first glance, the negative sign on the coefficient is surprising. The IRR reports that an increase in one Shorea robusta tree in the closest neighboring forest plot will result in nearly 15 percent decline in the number of Shorea robusta trees in the plot of interest. The influence could be explained by the natural strategy Shorea robusta trees take to replicate. The helicopter-like wings attached to the seed have developed so that they fly a good distance away from the parent tree. Winds take seeds away in a particular direction and several may take hold resulting in the natural clumped pattern exhibited by the Kaswang Forest.
Finally, of the other two location parameters in the model, the X and Y GIS coordinates, only X was found to be significant. The interpretation of this requires a review of Map 5.1. The X and Y coordinates reflect locations on this map, or grid. The Y coordinates follow the north-south axis, and the X coordinates follow the east-west axis. The origin is located in the center of the map-somewhere near the river convergence. The negative sign on the X coefficient suggests that as we move in an easterly direction, the trend in the number of Shorea robusta individuals decline. While the coefficient is small, it emerges as the most significant variable in the model.
Let us now return to the two research questions presented in the Introduction. The first question asks whether the spatial pattern of Shorea robusta differs in the two forests from what we found in the relatively undisturbed setting of the Kaswang Forest. The evidence provided in Table 5.2 strongly suggests that the spatial pattern of Shorea robusta within these two forests is different: the species does not follow a clumped pattern. This suggests that over space, Shorea robusta exhibits depletion in both the Sugabhanjyang and Latauli Forests.
The MLE regression location variables also support this conclusion. If the Shorea robusta individuals in these two forests follow a clustered or the negative binomial distribution as they should in an undisturbed setting, these location variables should not be significant predictors in the model. The distributional assumption used by MLE should account for the expected clumping pattern. Since both the X coordinate and elevation are found to be significant-and are, in fact, the two most highly significant variables in the model -they suggest that the spatial distribution of the Shorea robusta species in these forests follow a pattern different from what is expected in a natural setting. In short, the answer to research question one is: the data demonstrate that the natural spatial pattern of Shorea robusta has been disturbed in these two forests.
To answer research question two -what accounts for the distributional difference-I again turn to the location coefficients of the model. The elevation coefficient is highly significant and positive. This means that as one moves higher in elevation, the number of Shorea robusta species increase. What is interesting about this finding is that Shorea robusta are found in the lowest elevations in Nepal up to a maximum height of 1200 meters (Storrs and Storrs, 1990). The highest elevation in this study was approximately 800 meters. For this reason, in an undisturbed setting, I would not expect elevation to have any influence in the growth of this species after controlling for the clumped distribution of the species. I confirmed this expectation by running a similar regression on the data from the reference Kaswang Forest. In that instance, elevation is not found to be significant. Yet for the Sugabhanjyang and Latauli data, elevation is the second most significant influential variable in the model. This finding appears to support optimal foraging theory. Harvesting timber and carrying it back home from high elevations located far from the villages is both timeconsuming and labor-intensive. The elevation coefficient suggests that villagers prefer to harvest in plot areas that are found in lower elevations and closer to their homes.
The significance of the X coordinate is by far the most interesting finding. The negative coefficient of the X coordinate suggests that the trend in the count of Shorea robusta species diminishes as one moves east from the center of Map 5.1. The Latauli forest plots exhibit fewer numbers of the species. A number of alternative explanations exist that could account for this finding.
The first possible explanation for Latauli's depletion is that population in the Latauli area is greater than in the Sugabhanjyang area. However, IFRI data on village population cross-checked with 1990 census data both reveal that the population is greater in the western villages-the villages adjacent to Sugabhanjyang. The population data alone would suggest that Sugabhanjyang's Forest should exhibit less quantities of Shorea robusta-but it doesn't. Hence, a population explanation isn't supported.
The second possible explanation for the trend of depletion towards the Latauli Forest is an optimal foraging argument: that the Latauli plots are closer to the villages than Sugabhanjyang plots and therefore are more depleted. Just a glance at Map 5.1 proves this argument to be false. If anything, some Latauli plots are farther from villages.
The most convincing explanation for the Latauli depletion finding returns us to our earlier discussion of DFO monitoring and social norms that exist in the Shaktikhor area. My earlier description on DFO rules revealed that three formal rules exist that are applicable to all villagers in the region. However, the monitoring mechanisms are much stronger in the west along the road that follows the river just below and on the same side as the Sugabhanjyang Forest. This monitoring pressure in the west establishes an added incentive to harvest away from the road in the eastern Latauli Forest.
In addition, discussions with villagers on harvesting location preferences reveal the existence of social norms that also influence harvesting behavior. The villagers from lower castes, who reside in villages in the east, report that they only harvest in the nearby Latauli Forest. The higher-caste people however, living in the villages on the western side of Map 5.1, report that they harvest from both their nearby Sugabhanjyang Forest as well as the more distant Latauli Forest. These highercaste villagers actually break the first formal DFO rule when they forage in Latauli, for they cross a VDU boundary. The poorer villagers residing in the eastern side of Map 5.1 could make a legitimate complaint to the DFO if they desired, yet they do not. No conflicts have been reported, and both groups confirm these harvesting practices. Consequently, the best explanation for the diminishing trend of Shorea robusta towards Latauli is the combination of harvesting altered by more effective monitoring in the western side of Map 5.1 coupled with existing social norms between communities of differing caste structures.
While ecologists and biologists have made tremendous advances in the study of the spatial distribution of various plant species, to my knowledge, this is the first analysis of its kind that applies recent technological advances of GPS, GIS, and maximum likelihood estimation to this effort. The inclusion of a spatial influential variable in the regression model provides an alternative methodology for identifying early forest depletion when longitudinal data is nonexistent. The findings support my earlier claim that a plot level of analysis may reveal findings that would not be discovered at the forest level of analysis.
This study may also be the first of its kind to apply an institutional analysis to the study of the distribution of a particular species over space. It provides a new technique for researchers utilizing IFRI data to identify the influence of institutions on foraging behavior. While the study site was limited in terms of the institutional arrangements that could be modeled, this effort does show that a spatial analysis of IFRI forest plot data can be quite revealing in identifying how humans react to both written and unwritten rules and social norms. The location information in the regression model provides evidence that social norms produce resource inequities.
Let me conclude with three lessons to improve the methods utilized in this research. First, the researcher would be wise to document carefully location variables such as the distance of plots to nearby trails. This data, which was not collected in this study, may provide additional explanatory power in the model. Optimal foraging theory suggests that plots located closer to trails will be subject to greater harvesting if effective institutions have not been established to prohibit this activity. This additional information, added to the model, could assist us in understanding the influence of various institutional arrangements and would capture another spatial component of optimal foraging theory. Second, plot sampling should take into account the location of trails. In this study, the plots collected may have been biased toward a depleted state due to their close proximity to trails. An effective sampling strategy should select plots that are both near and distant from forest trails and document this distance accurately. Third, for quality regression results, plot locations should be meticulously recorded through the use of topographic maps and a carefully recorded traverse through the forest. If available GPS recordings are taken, these points should be differentially corrected to ensure their accuracy.46 Location errors inherent in one GPS receiver acting alone, or in careless recording on topographic maps, could cause significant problems for this kind of analysis.
Aldhous, P. 1993. `Tropical. Deforestation: Not Just a Problem in Amazonia." Science 259 (March 5): 1,390.
Ascher, W. 1995. Communities and Sustainable Forestry in Developing Countries. San Francisco, CA: ICS Press.
August, P., J. Michaud, C. Labash, and C. Smith. 1994. "GPS for Environmental Applications: Accuracy and Precision of Locational Data." Photogrammetric Engineering and Remote Sensing 60(1):41-45.
Feit, H. A. 1973. "The Ethno-ecology of the Waswanipi Uree, or How Hunters Can Handle Their Resources." In Cultural Ecology, ed. B. Cox, 115-25. Toronto: McUlelland and Stewart.
Hayden, B. 1981. "Subsistence and Ecological Adaptations of Modern HunterGatherers." In Omnivorous Primates, ed. R.S.O. Harding and G. Teleki, 344-421. New York: Columbia University Press.
IFRI (International Forestry Resources and Institutions). 1994. IFRI Data Collection Forms of the Shaktikhor Site. Bloomington: Indiana University, Workshop in Political Theory and Policy Analysis.
King, G. 1989. "Variance Specification in Event Count Models: From Restrictive Assumptions to a Generalized Estimator." American Journal of Political Science 33(3):762-84.
_____1990. Unifying Political Methodology: The Likelihood Theory of Statistical Inference. Cambridge: Cambridge University Press.
Kozlowski, T. T., P. J. Kraemer, and S. G. Pallardy. 1991. The Physiological Ecology of Woody Plants. San Diego, CA: Academic Press.
Lovejoy, T. E. 1980. "A Projection of Species Extinctions." In The Global 2000 Report to the President: Entering the 21st Century, ed. G.O. Barney (Study Director), 328-31. Washington, DC: Council on Environmental Quality, U.S. Government Printing Office.
Ludwig, J. A., and J. F. Reynolds. 1988. Statistical Ecology. New York: Wiley.
McKean, Margaret A. 1992. "Management of Traditional Common Lands (Iriaichi) in Japan." In Making the Commons Work: Theory, Practice, and Policy, ed. Daniel W. Bromley et al., 63-98. San Francisco, CA: ICS Press.
Myers, N. 1988. "Tropical Forests and Their Species: Going, Going..." In Biodiversity, ed. E.O. Wilson and F.M. Peter, 28-35. Washington, DC: National Academy Press.
Norton, B. J., ed. 1986. The Preservation of Species. Princeton, NJ: Princeton University Press.
Ostrom, Elinor. 1990. Governing the Commons: The Evolution of Institutions for Collective Action. New York: Cambridge University Press.
Ostrom, Elinor, et al. 1994. IFRI Coding Forms. Bloomington: Indiana University, Workshop in Political Theory and Policy Analysis.
Pardo, R. 1993. "Back to the Future: Nepal's New Forestry Legislation." Journal of Forestry 91:22-26.
Reid, W. V., and K. R. Miller. 1989. Keeping Options Alive: The Scientific Basis for Conserving Biodiversity. Washington, DC: World Resources Institute.
Repetto, R. 1988. The Forest for the Trees? Government Policies and the Misuse of Forest Resources. Washington, DC: World Resources Institute.
Richards, J. F., and R. P. Tucker, eds. 1988. World Deforestation in the Twentieth Century. Durham, NC: Duke University Press.
Shrestha, R. 1996. Personal communication, June.
Smith, E. A. 1983. "Anthropological Applications of Optimal Foraging Theory: A Uritical Review." Current Anthropology 24(5):625-52.
Spurn, S. H., and B. V. Barnes. 1992. Forest Ecology. Malabar, FL: Krieger.
Stephens, D. W., and J. R. Krebs. 1986. Foraging Theory. Princeton, NJ: Princeton University Press.
Storrs, A., and J. Storrs. 1990. Trees and Shrubs of Nepal and the Himalayas. Kathmandu: Pilgrims Book House.
Task Force on Global Biodiversity, Committee on International Science. 1989. Loss of Biological Diversity: A Global Crisis Requiring International Solutions. Washington, DC: National Science Board.
Thomson, James T., David Feeny, and Ronald J. Oakerson. 1992. "Institutional Dynamics: The Evolution and Dissolution of Common-Property Resource Management." In Making the Commons Work: Theory, Practice, and Policy, ed. Daniel W. Bromley et al., 129-60. San Francisco, CA: ICS Press.
Winterhalder, B. 1993. "Work, Resources and Population in Foraging Societies." Man 28:321-40.
30 Species "dominance", "frequency", "density, " and "importance value" are commonly used species abundance indicators. The dominance of a particular species is a measurement of the biomass that the species contributes to a forested area. The frequency of a species describes how widely it is distributed within a forested area. The density of a particular species is a count of the number of individuals that are present within the area sampled. Finally, the importance value combines these three indicators to provide a general indicator of abundance of the species.
31 An underlying argument made in this chapter is that more attention should be given to the analysis of important forest species in our studies of deforestation and biodiversity. The term "important species" is used to describe forest species that are found to be particularly useful by villagers for their day-to-day subsistence needs. Only by understanding what drives these species to be harvested or conserved will we fully understand why forest change has or has not occurred.
32 The term "space" is used to describe the forested area or landscape of study. A "spatial" analysis investigates relationships between objects of interest exhibited over this landscape.
33 This example is based on optimal foraging theory, which is described in further detail in a later section.
34 The term "natural, undisturbed setting" is used to describe a setting in which the important product species is not disturbed by human activity.
35 A VDC is the name given to the smallest political-administrative unit in the Nepali governmental system.
36 The research team is under the direction of Mr Rajendra Shrestha, Director of the Nepal Forestry Resources and Institutions Consortium in Kathmandu, Nepal.
37 The IFRI program defines a forest as an area larger than .5 hectares that is utilized by at least three households and that is governed by a similar legal structure (Ostrom et al., 1994).
38 Human stresses could be considered biotic variables (e.g., see Kozlowski, Kraemer, and Pallardy, 1991), but because of the importance of the human pressures in this study, I treat it as a separate category.
39 A winged seed.
40 GPS is a relatively new technology that uses a hand-held receiver to collect longitude, latitude, and altitude positions in the field. These receivers hold conversations with GPS satellites overhead and use trigonometry to calculate positions on the earth. Using a technique called differential correction, these recordings are accurate up to one meter Differential correction was used on this data.
One rather exciting side note should be mentioned here on the use of GPS for environmental research. During this field visit, we mapped a land-use boundary using GPS equipment. This effort was unique in that the boundary used to exist and no longer does. We followed a knowledgeable villager and mapped the area from his memory. GPS appears to provide researchers with a mechanism that can collect pail spatial relationships very accurately from the minds of villagers. It extends traditional participatory research techniques.
41 For example, Smith (1983: 632) describes the role that "exclusive control" plays in the conservation of natural resources. Feit (1973) describes rotational hunting by the Waswanipi Cree people as a method in which the size of animal population can be controlled.
42 Future studies that undertake a similar analytic method could easily establish variables to capture other institutional differences in forest plots. For example, if forest management units exist within the forest-areas within which harvesting of particular species are prohibited-a dummy independent variable could be specified to designate plots subject to this regulation.
43 King (1990) also argues that with the computational power available in today's personal computers, researchers should now move to more sophisticated regression models that are more true to our theoretical understanding of the real world instead of using traditional techniques that were developed for computational simplicity. In addition, models that violate the OLS assumption of normality force the researcher to implement statistical tricks to ensure that the residuals behave properly. This research avoids having to use statistical tricks by investigating and specifying the correct distributional assumption and then modeling it using MLE.
44 For a discussion of maximum likelihood event count models, see King (1989).
45 Literature describing the natural distribution of the Shorea robusta species was not available.
46 For a brief discussion on GPS accuracy, see August et al. (1994).
