![]()
![]()
Nous nous proposons ici d'examiner les connaissances techniques locales (CTL) de l'agriculteur traditionnel pratiquant la culture sur brûlis, et la façon dont elles sont appliquées à la gestion des ressources naturelles dans la zone tropicale humide. Après avoir passé en revue les grandes caractéristiques du milieu tropical humide et défini les problèmes que pose la gestion des ressources naturelles dans cette zone climatique, nous analyserons l'agriculture itinérante en tant que stratégie de gestion des ressources naturelles tropicales. Des exemples empruntés à trois grandes régions tropicales humides - le Bassin de l'Amazone, l'Asie du Sud-Est et l'Afrique centrale - viendront illustrer les pratiques d'agriculture itinérante en tant que phénomènes d'adaptation à l'environnement local, social et physique. En ce qui concerne l'Amazonie et l'Asie du Sud-est, les exemples se rapporteront à des minorités tribales qui ont, dans l'ensemble, très bien su utiliser et entretenir la forêt tropicale. Les exemples empruntés à l'Afrique montreront comment l'agriculteur pratiquant la culture sur brûlis s'adapte à un environnement moins stable, et comment se produit l'intensification des pratiques agricoles.
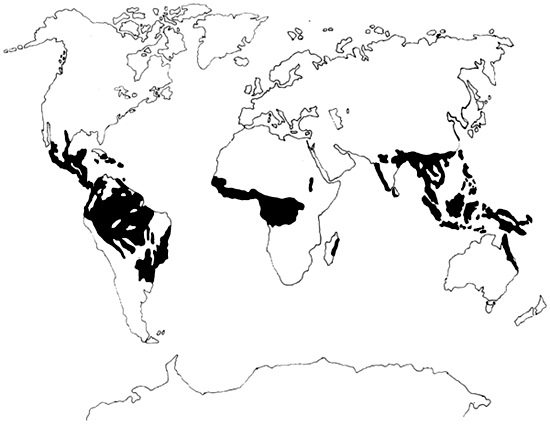
La déforestation de la zone tropicale préoccupe de plus en plus vivement la communauté internationale soucieuse de l'environnement. Les projections actuelles en cas de poursuite de la déforestation à grande échelle de la zone tropicale font redouter des phénomènes de crues, de sécheresse et d'érosion qui rendraient d'immenses régions impropres à l'agriculture. Certains travaux actuels sur les effets possibles, à l'échelle mondiale, du déboisement de la zone tropicale, laissent prévoir un réchauffement de la planète (effet de serre). Alors que l'on voyait autrefois dans les forêts tropicales une ressources naturelle devant être gérée à l'échelon national, on a aujourd'hui tendance à les considérer de plus en plus comme une ressource mondiale dont la gestion est l'objet de préoccupations au niveau international. Dans cette perspective, compte tenu des projections peu encourageantes que nous venons d'évoquer, on peut se demander ce qui a été fait pour protéger les forêts, qui détruit la forêt, et pourquoi il n'est pas mis fin aux abus?
Dans le passé (et dans certains cas aujourd'hui encore) les cultivateurs itinérants ont été les premiers accusés de la déforestation des tropiques. Des gouvernement et des organisations internationales, qui voyaient en eux les responsables de la destruction inconsidérée des ressources naturelles des nations, ont tenté de mettre fin à leurs pratiques. Or il était facile de promulguer des lois interdisant de couper et de brûler la forêt, mais mettre un terme à l'agriculture itinérante l'était moins. L'agriculture itinérante est une réalité, et continuera de l'être dans le futur.
Des études récentes ont montré que les principaux fautifs n'étaient pas ceux que l'on croyait. Loin de détruire la forêt à leur guise en ouvrant des clairières pour leurs cultures, puis en les abandonnant pour en ouvrir d'autres, bien des agriculteurs itinérants contribuent activement à sa régénération. L'agriculture itinérante est un système complexe qui, dans certaines conditions, est bien adapté aux contraintes du milieu tropical. II ne s'agit point d'une pratique primitive, et elle n'est pas nécessairement destructrice. Elle suppose au contraire une connaissance intime du milieu tropical et un grand savoir-faire pour porter ses fruits.
Ce point de vue nouveau sur l'agriculture itinérante a été renforcé par l'échec de certains projets de développement agricole sous les tropiques. Comme nous le verrons plus loin, les régions tropicales présentent un environnement peu propice à l'intensification de la production. Nombreux sont les projets qui ont échoué, laissant dans bien des cas derrière eux des formations herbacées, là où la forêt se dressait quelques années auparavant. Pourtant les agriculteurs itinérants, dans la même région, défrichaient et brûlaient la forêt, puis semaient et récoltaient, et la forêt revenait d'elle-même. Pourquoi les projets techniquement élaborés créeraient-ils des «déserts verts» et le cultivateur itinérant, pour primitif qu'il soit, régénérerait-il la forêt? En d'autres termes: que savent les cultivateurs itinérants, que font-ils, et pourquoi réussissent-ils sous les tropiques, quand d'autres approches échouent?
Dans ce qui suit, nous appellerons connaissances techniques locales (CTL) les connaissances pratiques de l'environnement et les stratégies en matière d'approvisionnement fondées sur une expérience intime accumulée au fil de maintes générations (Bodley 1976:48). Lorsque l'on étudie les connaissances techniques locales des agriculteurs itinérants, il faut avant tout recueillir des données de base sur «les ressources de l'environnement: végétaux, animaux, types de terres, sols, eau et cultures» (Knight 1980:222). Mais, quoique nécessaires, l'inventaire ethno-botanique et la classification des sols ne suffisent pas. Découvrir ce que le cultivateur itinérant connaît de l'environnement est certes important, mais ce qui compte surtout, c'est la façon dont ces connaissances sont mises en pratique. Compte tenu de sa connaissance et de sa perception du milieu, de la gamme de cultures possibles, de la disponibilité de terre et de main-d'œuvre, que fait effectivement l'agriculteur? II faut ici dépasser la simple description et classification des connaissances et tenter de comprendre comment celles-ci sont appliquées par l'agriculteur pour élaborer des stratégies en matière d'approvisionnement assurant sa sécurité alimentaire.
La connaissance que l'agriculteur pratiquant la culture sur brûlis a de l'environnement lui permet tout d'abord de prendre des décisions sur ce qu'il faut faire et quand il faut le faire. C'est ainsi que ses connaissances sont mises à l'épreuve: si la pratique les confirme, elles rejoignent le bagage de connaissances actives; si elles échouent, elles seront reléguées dans la catégorie de ce qui ne vaut plus, et abandonnées. Pourtant la «séquence de prise de décisions» repose sur d'autres facteurs que la connaissance de l'environnement: certaines contraintes ou situations de fait limitent le choix. Ces contraintes peuvent être d'ordre social, culturel ou environnemental (Ellen 1982). Certaines d'entre elles peuvent être de courte durée (situation conjugale, présence de jeunes enfants, maladie), d'autres étant constantes et relativement impératives (facteurs climatiques n'autorisant pas certaines cultures). Or c'est en appliquant les connaissances techniques locales et en s'accommodant de ces contraintes que l'agriculteur pratiquant la culture sur brûlis prend ses décisions et crée un système viable de production vivrière.
Concevoir ainsi l'agriculteur comme un décideur qui fait le compte de ses «ressources biologiques et économiques» et prend des décisions «en vue d'obtenir une production agricole et d'entretenir la fertilité du sol» conforte l'opinion actuelle selon laquelle l'agro-écosystème (le système agricole en tant qu'élément de l'écosystème «naturel» plus grand) est dynamique et synergique, plutôt que statique (Benneh 1972:245). La théorie de l'agro-écosystème considère l'agriculteur comme un participant actif, sa culture s'étant édifiée en symbiose avec l'évolution de l'environnement pour produire un système viable d'approvisionnement vivrier (Gliessman 1985:56). Les changements induits par les interactions entre l'homme, sa culture et l'écosystème entraînent à leur tour d'autres changements au fur et à mesure que de nouvelles décisions sont prises, après nouvelle évaluation des ressources. Cette dynamique, avec ses mécanismes complexes de rétroaction, permet de mieux comprendre comment l'agriculteur pratiquant la culture sur brûlis intègre le milieu naturel et le système agricole pour obtenir une production culturale durable (Gladwin 1983, Olafson 1983, Warner 1981, Benneh 1972).

Caféiculture et riziculture sur brûlis, en Thaïlande.
Autrefois pratiquée dans les forêts tempérées, l'agriculture itinérante est un agro-écosystème que l'on rencontre principalement aujourd'hui en milieu tropical humide. On définit la zone tropicale humide comme une région présentant les caractéristiques suivantes:
La zone ainsi définie couvre pratiquement 2500 millions d'hectares, répartis entre quatre régions: l'Afrique, l'Amérique du sud, l'Amérique centrale et l'Asie du Sud-Est (voir Tableau 1). En Afrique et en Amérique tropicale, l'écozone tropicale humide est principalement concentrée dans deux bassin hydrographiques. En Amérique tropicale, le bassin amazonien compte pour 75% de la zone humide. L'Amazonie est si grande qu'à elle seule elle représente plus de 40% de la zone tropicale humide mondiale (Sanchez 1987). En Asie du Sud-Est, la zone tropicale humide se partage entre la masse continentale et les îles équatoriales, à l'exclusion des zones de haute altitude.
Si toutes les régions mentionnées présentent les caractéristiques générales de la zone tropicale humide, les précipitations varient de l'une à l'autre et à l'intérieur de chacune d'entre elles. C'est en Amérique du Sud que les pluies sont les plus régulières, avec les plus faibles variations mensuelles, alors que dans pratiquement toute l'Afrique tropicale, il existe une saison sèche marquée, durant un à deux mois, au cours de laquelle la pluviométrie est inférieure à 100 millimètres (Richards 1973).
Forêt: la végétation naturelle de la zone tropicale humide est la forêt (Richards 1977: Hadly et Lanly 1983). II existe deux types principaux de forêts: la forêt dense, et la forêt claire (Hadly et Lanly 1983). La forêt dense est présente là où les précipitations annuelles moyennes sont supérieures à 1600 millimètres. Elle présente une voûte continue, est étagée, et la végétation de sous-bois est normalement abondante. Selon les régions, elle peut être composée de feuillus, de conifères ou de bambous. La composition de sa flore est variable, mais chaque type est adapté à des conditions climatiques analogues: précipitations abondantes et températures élevées (Hadly et Lanly 1983; Richards 1973).
Là où la pluviométrie est comprise entre 1200 et 1600 millimètres, le couvert naturel peut être plus ou moins dense, selon la durée de la saison sèche, les sols, etc. (OTA 1984). On trouve des forêts claires lorsque la pluviométrie est comprise entre 900 et 1200 millimètres, dans les régions plus sèches que celles qui portent une forêt dense. La forêt claire, ou ouverte, est du type mixte, alliant les arbres et les herbages. La voûte forestière est discontinue, mais couvre plus de 10% de la surface du sol.
Les forêts, denses et claires, sont inégalement réparties dans les régions tropicales. En Afrique tropicale, on ne trouve que 18% des forêts tropicales denses, mais 66% de la forêt claire mondiale. Celle-ci est caractéristique des lisières les plus sèches du Bassin du Congo et de l'Afrique de l'Est. Cinquante-sept pour cent de la forêt tropicale dense mondiale se trouve en Amérique tropicale, en majeure partie dans le Bassin amazonien, et 25% en Asie - près de la moitié étant concentrée en Indonésie (Hadly et Lanly 1983: OTA 1984).
C'est la forêt tropicale dense qui est, biologiquement, la plus complexe et présente la plus forte diversité génétique. C'est précisément cette forêt qui est victime du déboisement. L'homme, notamment depuis que l'agriculture a été adoptée comme système de subsistance, est responsable de la transformation en un semi-désert d'un milliard d'hectares, selon les estimations, de zone tropicale humide, soit une superficie équivalente à celle du Bassin amazonien tout entier (Benneh et al. 1977). Le rythme de la déforestation s'est accéléré depuis 20 à 30 ans, à mesure que l'élevage extensif, les plantations et l'exploitation forestière gagnaient, et que des agriculteurs migrants s'installaient en nombre croissant dans les forêt tropicales (Richards 1977).
Tableau 1. Etendue de la zone tropicale humide chaude (millions d'hectares)
| Région | Afrique | Amérique du sud | Amérique centrale | Asie du Sud-Est | Total |
|---|---|---|---|---|---|
| Etendue de la zone tropicale humide chaude | 911,7 | 1001,5 | 76,3 | 491,8 | 2481,3 |
| Pourcentage de la superficie totale par région | 31,7 | 56,5 | 28,1 | 54,8 | 38,2 |
Source: Ofori, Higgins et Purnell 1986 (citant FAO 1980; 1981; 1982)
Laissés intacts, les écosystèmes forestiers tropicaux sont stables. Cette stabilité résulte de leur capacité de «s'accommoder des accidents, climatiques et autres, de l'environnement naturel» (Richards 1977:230). Diverses caractéristiques de la forêt tropicale concourent à cela:
La forêt tropicale humide est riche en espèces végétales et animales. C'est ce degré élevé de diversité génétique qui assure la stabilité de l'écosystème forestier.
Les forêts tropicales sont les écosystèmes terrestres les plus complexes qui existent (Connell 1978). Végétaux et animaux sont intimement liés au sein de l'écosystème forestier tropical. En ce qui concerne la dissémination des graines et la pollinisation, les animaux assurent la même fonction que le vent dans la forêt tempérée (Hadly et Lanly 1983:5). Etant donné que la forêt tropicale présente des espèces beaucoup plus diversifiées, et que les animaux se déplacent dans un petit périmètre, la régénération et l'entretien de la diversité locale sont assurés.
Les sols tropicaux étant en général pauvres en nutriments, l'écosystème forestier tropical repose sur un cycle des nutriments, auto-entretenu, quasiment fermé. Le cycle nutritif se caractérise par le mouvement des nutriments au sein de la biomasse, qui a une fonction de stockage végétatif. Soixante-cinq à 85% du système racinaire ne descendant pas au-dessous de la couche superficielle du sol, la forêt elle-même fonctionne comme une sorte d'éponge géante, qui absorberait et recyclerait les nutriments (Hadly et Lanly 1983; Uhl 1983; Moran 1981).

L'écosystème forestier tropical repose sur un cycle des nutriments, auto-entretenu, quasiment fermé
Les études consacrées à l'Amazonie ont montré l'importance du tapis racinaire des arbres dans le cycle des nutriments. Cette masse fibreuse, composée des radicules des arbres entremêlées de matière organique et de champignons mycorhiziens, recouvre le sol et constitute la litière forestière. Lorsque les feuilles, les ramilles, voire les arbres entiers tombent sur ce tapis et commencent à se décomposer, cette couche superficielle absorbe les nutriments dissous avant qu'ils ne puissent être lessivés vers le sol (Stark et Jordan 1978). Comme 10 à 20% de la biomasse totale meurt et tombe au sol chaque année, la quantité de nutriments recyclés par le système est importante (Moran 1981).
Ce système est d'une efficacité telle que «la concentration de certains nutriments dans les cours d'eau qui drainent la forêt est en fait inférieure à leur concentration dans les pluies qui l'arrosent» (Uhl 1983:70). Dans la forêt, non seulement les arbres mais aussi d'autres plantes ont acquis une moindre dépendance vis-à-vis du sol—les épiphytes, qui vivent sur les feuilles des arbres, sont capables de tirer les nutriments de l'eau de pluie et de fixer l'azote de l'air (Uhl 1983). La forêt est donc un écosystème qui, une fois constitué, s'auto-entretient pour autant que la pluie continue à l'arroser et qu'on le laisse intact.
Pourtant la forêt, aussi stable soit-elle, n'est pas un milieu statique. La chute naturelle des arbres fait partie du processus d'auto-entretien de la forêt. La forêt tropicale n'est pas une forêt vieillie, elle est un lieu de changement et de renouvellement constants, grâce au vent et à la chute des arbres. L'arbre qui tombe ouvre un interstice dans la voûte, et une tache de lumière solaire peut ainsi atteindre le tapis forestier. Plus grande est cette ouverture, plus étendu sera le micro-climat ainsi créé, et plus variées seront les espèces végétales qui pourront se développer dans le créneau ainsi ouvert. Dans un écosystème où les nutriments sont emmagasinés dans la biomasse, la chute d'un arbre par demi-hectare et par an assure un surcroît notable de nutriments disponibles (Hadly et Lanly 1983; Uhl 1983, Hartshorn 1978; Whitmore 1978).
La fréquence élevée des chutes d'arbres, notamment dans les régions tropicales où sévissent de fortes tempêtes ou des cyclones, empêche la plupart des arbres de réaliser leur plein potentiel de croissance, en taille ou en âge. Les espèces arborées pionnières de la succession requièrent des ouvertures dans le couvert, car elles ne pourraient pas se développer sans lumière solaire et sans l'afflux de nutriments que déclenche la chute d'un arbre. Le type d'espèces qui prennent la succession dans l'ouverture est quant à lui déterminé par les relations qui règnent localement entre végétaux et herbivores. Ces facteurs génèrent une mosaïque d'ouvertures dans la voûte et déterminent les divers stades de croissance dans le sous-étage qui donnent à la forêt tropicale une diversité unique en végétaux et en animaux. C'est une forêt dynamique, caractérisée par la croissance rapide des espèces pionnières et la croissance relativement lente des essences forestières de climax; la forêt présente ainsi une série de taches, correspondant à divers stades de recrû, au sein d'un système en équilibre, globalement stable. (Hadly et Lanly 1983; Hartshorn 1978; Whitmore 1978).
Mais une telle stabilité ne peut se manifester que dans le cadre de processus naturels de renouvellement. L'homme représente un véritable danger pour les forêts tropicales, surtout lorsqu'il y pénètre non pas armé d'une hache, mais d'une tronçonneuse, ou assis sur un bulldozer. Les facteurs mêmes - diversité, complexité et cycle fermé des nutriments-qui assurent la pérennité de l'écosystème forestier tropical hors de toute intervention extérieure deviennent des facteurs de fragilité dès qu'intervient l'homme. La forêt humide, en raison du degré élevé de spécialisation de chaque espèce, a une faible capacité de récupération quand elle est bouleversée par l'homme sur une grande échelle (Goudie 1984; Hill 1975). C'est la complexité même de l'écosystème forestier tropical — qui assure sa stabilité à l'état naturel — qui le rend aussi vulnérable aux interventions de l'homme.
Cette vulnérabilité est aggravée par la manière dont se produit la repousse de la végétation. La reconstitution de la forêt, lorsqu'une ouverture s'est produite naturellement avec la chute d'un arbre ou après ouverture d'une petite éclaircie (d'une superficie de moins de trois hectares), se produit selon quatre axes principaux:

Les graines d'essences forestières peuvent être introduites dans la trouée par des mammifères, des oiseaux ou des chauves-souris.
Ces modes de reprise de la végétation sont efficaces lorsque l'ouverture est petite, mais leurs limites apparaissent lorsque la superficie défrichée est grande, qu'elle résulte de l'exploitation forestière classique ou du passage de bulldozers. Dans ces cas-là, les jeunes plants ne subsistent qu'au périmètre de l'éclaircie, et il ne se trouve pas d'arbres dans celle-ci pour rejeter; les graines en dormance sont emportées avec la litière végétale, et le réensemencement par la faune devient difficile, car la surface dénudée est trop grande pour attirer les oiseaux et les chauves-souris, ou pour que des mammifères s'y sentent suffisamment en sécurité pour y pénétrer (Jordan 1985).
A cela vient s'ajouter l'effet du cycle des nutriments dans la forêt tropicale. Les nutriments étant stockés dans la biomasse, une fois la forêt enlevée, ils ne sont plus assez abondants pour soutenir la reconstitution de la végétation. Dès qu'il n'est plus protégé par le couvert forestier, le sol est emporté par les grosses pluies, puis durci par l'exposition directe au soleil. La taille de l'ouverture, l'enlèvement de la couche superficielle du sol, et l'exposition à la pluie et au soleil se combinent pour ralentir, de façon spectaculaire, la réinstallation de la forêt. Il pourrait falloir jusqu'à mille ans pour qu'une ouverture de 15 hectares, pratiquée au bulldozer, et dégarnie de toute végétation, soit de nouveau couverte par la forêt (Uhl 1983).
Sols: Quoique les sols de la zone tropicale humide puissent présenter une grande diversité, ils manquent pour la plupart de nutriments (Jordan 1985). Dans la zone tropicale humide d'Afrique, en Asie du Sud-Est et en Amazonie, les problèmes posés par le déficit en phosphore, la toxicité due à l'aluminium, la faible résistance à la sécheresse, et la faible fertilité inhérente sont courants et bien connus (Sanchez 1987; Lal 1989; Moorman et Kang 1978). La pluviométrie est le facteur qui semble être à l'origine de la médiocrité des sols de la région, car dès que les précipitations dépassent 1000 millimètres, on constate que les sols sont habituellement acidifiés et lessivés (Sanchez 1987).
Les carences en nutriments présentées par les sols tropicaux sont le principal facteur limitant la productivité. Certes ces sols «vieillis, usés par les phénomènes atmosphériques et excessivement lessivés» portent une forêt tropicale humide, mais celle-ci ne dépend pas d'eux pour satisfaire ses besoins en nutriments (Lal 1987:16). Au contraire, l'écosystème forestier tropical s'abstrait du sol et crée son propre cycle de nutriments, qui repose sur sa propre biomasse. Contrairement aux zones tempérées où la taille des arbres de la forêt est en gros proportionnelle à la fertilité du sol, la taille des arbres de la forêt tropicale n'est pas représentative de la teneur en nutriments des sols sous-jacents (Jordan 1982; 1985). Les nutriments migrent des feuilles, des fûts tombés en chablis, etc., par les mycorhizes et les racines superficielles de la litière forestière pour se réintégrer à la biomasse, mais n'atteignent jamais le sol proprement dit (Beckerman 1987:64; Went et Stark 1968).
Une fois que la déforestation s'est produite et que le cycle des nutriments de l'écosystème forestier est rompu, le sol perd ses nutriments et sa structure physique est affaiblie. Même si la forêt tropicale n'était pas dépendante du sol pour son approvisionnement en nutriments, les racines des arbres exerçaient une fonction fixatrice et permettaient l'infiltration de l'eau, tandis que la litière forestière protégeait le substrat de la pluie (Goudie 1984). Une fois cette litière forestière enlevée, le sol devient sensible au phénomène de compactage, il perd ses propriétés de rétention de l'eau, et ne peut plus abriter une macro-faune importante (vers de terre et termites), qui lui apportait des nutriments et en améliorait la structure physique (Lal 1987). Avec la déforestation, la protection du sol assurée par la forêt disparaît. Les sites déboisés, surtout s'ils occupent plus de quelques hectares, subissent une érosion accélérée, grave le cas échéant, dès qu'ils sont exposés à de fortes pluies.
La vulnérabilité du sol à l'érosion est déterminée par la superficie de la zone défrichée et la méthode utilisée. Si l'éclaircie est petite, de superficie inférieure à 2 ou 3 hectares, et si elle reste entourée par la forêt, la végétation réapparaîtra rapidement, et la perte de sol par érosion sera minime. Si la superficie est plus grande, le sol s'appauvrira rapidement en nutriments et subira davantage les effets de l'érosion. Mais même une petite ouverture peut occasionner de forts écoulements superficiels et provoquer des phénomènes érosifs si elle est pratiquée selon des méthodes très perturbatrices.
Tableau 2. Effets des méthodes de déboisement sur l'écoulement de surface et l'érosion
| Mode de déboisement | Ecoulement superficiel (mm/an) | Erosion du sol (t/ha/an) |
|---|---|---|
| Défrichage traditionnel (Coupe sélective) | 3 | 0,01 |
| Manuel | 35 | 2,5 |
| Cisailles | 86 | 3,8 |
| Bélier d'abattage/rateau déracineur | 202 | 17,5 |
Source: Lal 1987

Défrichage d'une petite superficie par des moyens traditionnels au Honduras
Le défrichage de la forêt par les méthodes traditionnelles et manuelles entraîne une érosion du sol moins grave que le déboisement par des moyens mécaniques, notamment avec des béliers d'abattage (voir Tableau 2). La méthode de défrichage qui provoque le moins de ruissellement et d'érosion est la méthode traditionnelle, qui ne fait appel qu'à la machette et à la hache; la plus agressive est celle qui fait appel au bélier d'abattage et au râteau déracineur. Les taux d'érosion sont directement fonction de la végétation qui reste sur le site après défrichage. Les méthodes traditionnelles laissent en place les souches et ne touchent pas aux systèmes racinaires, tout en dérangeant relativement peu la litière forestière; si la pleine protection assurée par le couvert forestier a disparu, les racines continuent de maintenir le sol, et la litière fait écran à l'impact des gouttes de pluie. Les béliers d'abattage défrichent le terrain en faisant basculer les arbres et en arrachant les racines. Après défrichage, il ne reste qu'une étendue sans racines, pratiquement sans litière, et la surface du sol est profondément perturbée. Le défrichage avec les engins modernes provoque un ruissellement et une érosion graves, avec un ruissellement près de 70 fois supérieur et une perte de terre multipliée par un facteur de 1700 par rapport au défrichage traditionnel.
Les estimations du nombre effectif de cultivateurs itinérants vont de 250 millions (Myers 1986) à 300 millions (Russell 1988). Sur une population mondiale de 5 milliards de personnes, la manière dont 5% de la population assure sa subsistance pourrait sembler peu importante. Mais on ne saurait ignorer la distribution géographique des populations pratiquant l'agriculture itinérante et la grande superficie sur laquelle leurs systèmes agro-forestiers sont pratiqués. L'agriculture itinérante est la technique de gestion des sols tropicaux la plus répandue. Divers types d'agriculture itinérante sont actuellement pratiqués sur 30% des sols exploitables dans le monde (Hauck 1974, Sanchez 1976:346).
Il existe plusieurs définitions de l'agriculture itinérante. Le plus souvent, on appelle agriculture itinérante tout système agricole dans lequel les champs sont défrichés (habituellement par le feu) et cultivés pendant une période brève pour être ensuite mis en jachère (Conklin 1957). Avec l'apparition de la théorie des agro-écosystèmes, qui inscrit les systèmes agricoles dans un écosystème naturel plus grand, la définition de l'agriculture itinérante a été repensée. La théorie des agro-écosystèmes s'efforce d'intégrer «la multiplicité des facteurs entrant en jeu dans les systèmes culturaux» (Gliessman 1985:18). Tandis que de nombreuses études antérieures présentaient le système de culture sur brûlis comme essentiellement stable et répertoriaient ses caractéristiques, des travaux plus récents fondés sur la théorie des agro-écosystèmes inscrivent les systèmes de culture sur brûlis et de jachère dans une stratégie globale de subsistance, qui permet de répondre avec souplesse aux différentes contraintes s'exerçant sous l'effet de l'évolution du milieu social, économique ou naturel (Gliessman 1985, Altieri et al. 1973).
Traduisant cette vision dynamique, McGrath (1987:223) a décrit plus récemment l'agriculture itinérante comme «une stratégie de gestion des ressources selon laquelle on déplace les cultures pour exploiter le capital énergétique et nutritionnel du complexe naturel de végétation et de sol d'un nouvel emplacement». L'accent placé sur la stratégie et la dynamique de l'agro-écosystème fait de l'agriculture itinérante «un système de production agricole qui n'est ni statique, ni nécessairement stable», mais qui fait preuve de souplesse vis-à-vis des changements (McGrath 1987:223).
Considérer l'agriculture itinérante comme une stratégie caractérisée par sa flexibilité vis-à-vis du changement la replace dans le cadre des autres systèmes agricoles, qui peuvent s'en distinguer par la durée de la jachère, la durée de la période de culture, les techniques de gestion, etc.. Le passage d'un système de production à un autre se produit en réponse à la modification des conditions ambiantes (Beckerman 1987; Boserup 1965; Raintree et Warner 1986).
En tant que stratégie de subsistance, l'agriculture itinérante n'est guère appréciée par les pouvoirs publics et les organismes internationaux. On la considère couramment comme un gaspillage de ressources en terre et en hommes, et on l'accuse d'être un facteur majeur d'érosion et de dégradation des sols. Déboiser, pratiquer des cultures sur brûlis pendant un an ou deux, puis passer à un autre morceau de forêt peut effectivement ressembler à du gaspillage si l'on ne pense à la forêt qu'en fonction de la valeur du bois d'œuvre qu'elle contient (Grinnel 1977; Arca 1987). Mais la polémique est moins liée à l'abattage de la forêt, que les forestiers pratiquent en permanence, qu'au fait que l'on brûle les arbres. On se préoccupe moins de sauvegarder (ou de ne pas déranger) la forêt que de voir échapper des recettes potentielles. Les pouvoirs publics considèrent que l'écobuage pratiqué à l'échelon local (celui des petits agriculteurs) les dépossède de ressources nationales.
En Afrique, l'agriculture itinérante est pratiquée par les populations de toute la zone humide. Mais les longues périodes de jachère ont été progressivement remplacées par l'exploitation intensive de champs à proximité du lieu d'habitation, et par des assolements plus longs des parcelles éloignées (Chidumayo 1987; Getahun et al. 1982). Même si les pratiques effectives de gestion, les cultures pratiquées, etc., varient dans une certaine mesure, cette intensification de l'agriculture itinérante est un phénomène général dans toute la région.
Contrairement à l'Afrique sub-saharienne, où l'appartenance à un groupe tribal est de règle pour tous les individus, en Asie et en Amérique latine les cultivateurs itinérants pratiquant la jachère forestière appartiennent à des minorités ethniques qui ont chacune leur langue, leur religion, leurs valeurs et, dans certains cas, leurs propres formes d'agriculture. La perception qu'ont les pouvoirs publics de l'agriculture itinérante comme système d'utilisation des terres est étroitement liée au fait qu'elle est pratiquée par des populations culturellement marginales. Les cultivateurs itinérants sont considérés comme «primitifs» parce que leur culture matérielle est plus simple, ou tout simplement différente, et ainsi l'agriculture qu'ils pratiquent est elle aussi considérée «primitive», gaspillant semble-t-il des ressources qui seraient certainement mieux mises à profit à l'échelle de la nation.
Ce parti pris a fait obstacle à l'apparition d'une vision plus objective de l'agriculture itinérante dans bien des pays. Ainsi le système d'utilisation des terres est jugé en fonction de qui le pratique, plutôt que sur ses mérites et ses limites propres. De plus, en Asie comme en Amérique latine, la perception de l'agriculture itinérante est plus difficile du fait que non seulement les minorités tribales, ou les «indios» la pratiquent, mais aussi les paysans sans terre et les migrants qui colonisent des terres vierges. Dans le meilleur des cas, on reste indifférent à ce qui peut advenir des groupes appartenant aux échelons les plus bas de la société, sauf si l'on estime qu'ils s'approprient indûment les ressources nationales. Les paysans comme les minorités tribales pratiquent une agriculture itinérante, au sens large, mais leurs systèmes respectifs d'utilisation des terres sont fondamentalement différents.
Les groupes tribaux pratiquent en général la culture sur brûlis intégrale, à savoir un système d'utilisation des terres fondé sur «un mode de vie traditionnel, permanent, étendu à toute la communauté, largement autonome et sanctionné par un rituel». Lorsque ces agriculteurs pénètrent dans une zone nouvelle en tant que pionniers, ils défrichent chaque année des fractions notables de la végétation à son apogée. Une fois la communauté bien établie, l'éclaircie annuelle ne porte pratiquement plus sur la végétation de climax, et c'est alors la culture sur brûlis intégrale stabilisée qui est pratiquée (Conklin 1957:2, 3).
Les paysans, quant à eux, pratiquent la culture sur brûlis partielle, qui n'est pas fondée sur un mode de vie, mais traduit de «façon prédominante les seuls intérêts économiques des participants» (Conklin 1957:2). Ces paysans ont des liens socio-culturels étroits avec l'extérieur de la zone immédiate où ils pratiquent leurs cultures, et leurs objectifs en termes de propriété et de productivité sont différents de ceux qui pratiquent la culture sur brûlis intégrale. Au lieu d'appartenir à une communauté stable possédant des liens historiques et culturels avec la zone géographique, l'agriculteur pratiquant la culture sur brûlis partielle peut fort bien ne s'y trouver que pour une année ou deux. Ce mode de culture est donc essentiellement pratiqué par des agriculteurs cultivant des champs permanents, qui n'y ont recours qu'en complément de leurs cultures permanentes. Dans ces cas, l'agriculteur pratique une culture sur brûlis d'appoint. En Asie du Sud Est, il est fréquent que les parcelles permanentes soient dans les vallées, et les champs sur brûlis à flanc de colline. Un autre système de culture sur brûlis partielle est pratiqué lorsque le cultivateur migre dans la forêt. Souvent dépourvu de connaissances préalables en techniques de culture sur brûlis, cet agriculteur y consacre cependant tous ses efforts. Il s'agit alors d'une culture sur brûlis embryonnaire, mais dans la plupart des cas, l'agriculteur ne possède pas les connaissances voulues pour établir un système susceptible de durer (Conklin 1957:3).
De telles distinctions ont souvent été faites dans la documentation, mais l'on distingue cependant une tendance, surtout en Amérique du Sud, à confondre culture sur brûlis embryonnaire et pionnière. Plutôt que d'utiliser le terme pionnier dans son acception originelle (communauté tribale pratiquant le brûlis intégral et venant s'établir dans une zone nouvelle), l'expression «pionnière» est improprement utilisée pour décrire les modes de culture des paysans migrants qui pénètrent dans la forêt, y pratiquent des cultures sur brûlis, puis abandonnent le site ou vendent un champ dégradé, ou bien le convertissent à des cultures permanentes (UNESCO/PNUE 1978:324; Moran 1987). Selon les définitions originellement données par Conklin, ces paysans migrants ne pratiquent pas une culture sur brûlis de type pionnier, mais une culture sur brûlis embryonnaire qui se dégrade faute de connaissances sur l'écosystème forestier. Néanmoins, l'usage s'étant établi depuis quelques années, nous utiliserons dans la suite du présent document l'expression «pionnières» pour nous référer aux pratiques des nouveaux migrants, pour les distinguer de la culture sur brûlis intégrale pratiquée par les communautés autonomes bien établies.
Parmi ces millions de cultivateurs itinérants, combien pratiquent la culture sur brûlis pionnière, et combien l'intégrale? Bien des gouvernements, hélas, ne voient aucune différence entre ces deux catégories (la culture sur brûlis intégrale est aussi souvent qualifiée de traditionnelle). Comme ces deux systèmes ont des effets très différents sur l'environnement, il convient pourtant d'établir une distinction (Watters 1971). Lorsque la forêt tropicale est détruite, c'est en général l'agriculture pionnière, et non pas l'intégrale qui est en cause. Les migrants, avides de terres, sans tradition de culture sur brûlis intégrale, qui leur procurerait les connaissances voulues pour gérer l'écosystème forestier, pénètrent dans les zones forestières, les cultivent et les dégradent (Olafson 1981:3: voir aussi Moran 1987:227; Moran 1983; Watters 1971). Toute population qui réside dans une zone pendant une génération au moins possède une connaissance beaucoup plus précise du milieu local que le migrant à la recherche d'un terroir, lequel risque davantage de pratiquer un système pionnier, appliquant des méthodes agricoles de sa région d'origine plutôt que celles qui conviennent à la zone où il s'installe (Moran 1987:227).
![]()
![]()