3.3 Trees and shrubs as a source of livestock fodder
3.3.1
Trees and shrubs within pastoral systems
3.3.2
Fodder production and nutritive value
3.3.3
Improved use of tree fodder
Another important way in which trees and shrubs contribute to food security is by providing fodder for livestock. In some cases, trees are deliberately planted for their fodder and foliage is cut by hand for stall feeding to animals. More commonly, livestock is allowed to browse on the trees and shrubs growing naturally within grazing areas. Fodder from forest areas, whether browsed or harvested helps sustain livestock production and helps to ensure a year round supply of milk, blood and meat products.
3.3.1 Trees and shrubs within pastoral systems
Trees and shrubs are particularly important in pastoral production systems. Communities who gain their livelihood from herding animals depend for their survival on an intimate knowledge of their environment. Trees and shrubs are recognised as an essential component within these systems.
There are between 30 and 40 million pastoralists worldwide. Of these, 20-25 million are in Africa, especially in the dry sub-Saharan belt stretching from Mauritania to Ethiopia. The density of woody species within these areas and their relative importance as a source of animal feed is determined primarily by the availability of water. In the driest parts, woody vegetation is scarce and tends to be concentrated along drainage lines and in low lying land where underground water is available. As rainfall increases, woody species become more common.
The range of woody species that are used for animal fodder is extremely wide (Skerman, 1977; Felker and Bandurski, 1979). The fodder they provide, which is collectively termed browse, consists of a combination of leaves, small branches, seed pods and fruits. The importance of browse depends on the type of livestock. Camels and goats are great consumers of leaves and small branches of woody species, while cattle and sheep rely mainly on grasses and annual herbs (Lusigi, 1981).
In many pastoral areas, fodder from trees and shrubs is an indispensable part of livestock diets (le Houerou, 1986; Torres, 1983). This is particularly so during the dry season when the nutritional quality of the herbaceous layer is markedly reduced. At the beginning of the dry season rapid evapotranspiration takes place and the content of digestible protein and 8-carotene (necessary for the synthesis of Vitamin A) drops significantly. The dietary energy available also falls due to lignification and the increase in cellulose content at the expense of the more digestible hemicellulose. Animals eating only dry grass would suffer from malnutrition, both because of the shortage of energy and protein, and because of the lack of Vitamin A and essential minerals, especially phosphorus.
Herders recognise the critical role of trees and shrubs. In Sahelian West Africa, most herding groups are well aware of the causes and dangers of vitamin A deficiency, and whenever possible manage their herds so as to avoid pastures with no shrubs or trees.
In part of northern Senegal, it is estimated that during at least 6 months of the year the herbaceous vegetation is not adequate for livestock if used alone because of its high cellulose content and low nutritional quality. Pastoralists are only able to maintain their herds because of the availability of high quality supplements leaves, fruits and seed pods from trees and shrubs (Bille, 1977). As a fraction of total fodder intake during the dry season, browse can contribute as much as 30% of cattle's and 60% of goats' fodder.
3.3.2 Fodder production and nutritive value
A number of attempts have been made to measure fodder production from different components of dryland ecosystems (Trollope, 1981). In general while trees and shrubs are less prolific than annual herbs and grasses, the productivity of grasses tends to be extremely variable, mainly in response to fluctuations in rainfall between seasons, and from year to year. Production of browse from trees and shrubs is much more stable, and because of deeper root systems trees and shrubs are less susceptible to short-term localised fluctuations in rainfall.
The nutritive value of any fodder depends not only on its nutrient content but also on the amount consumed and assimilated by the animal. Although there are a wealth of data on the chemical composition of different types of browse, there is very little information on their effectiveness as animal feeds.
Protein content is one important variable, as this is the main limiting factor affecting liveweight gains of livestock in semiarid areas (Pratchett et al, 1977). Here, browse tends to have an advantage over grass. Comparisons of different types of fodder in the coastal and interior savanna of Ghana, for example, found that browse typically has 2 to 3 times as much protein as grasses, the exact figures varying between seasons.
Though it will generally be the case, fodder with a high protein content is not necessarily a good protein supplement. Measurements of protein digestibility show considerable variations between plant species; with Prosopis cineraria, for example, sheep are able to digest only 22% of the protein present, according to one set of measurements, compared to 83% for Atriplex nummularia. There are also differences between animals; goats are able to obtain more than twice as much protein from Ficus bengalensis than cattle (Torres, 1983). Thus, simply knowing the species which are commonly browsed and the nutrient composition of their fodder will not give an indication of their value as livestock fodder. The most important feature of forest fodder is that it is available in periods when other feed is unpalatable or non-existent.
3.3.3 Improved use of tree fodder
An increasing problem for pastoralists in many areas is the growing pressure on tree fodder resources. Overbrowsing leads to reduced regeneration of trees and shrubs, and carried to extremes will result in their gradual eradication. In parts of the Sahel this has been an important factor in the decline of Acacia seyal and A. senegal (le Houerou, 1986). similarly, in the central rangelands of Somalia, Yicib (Cordeauxia edulis), which is the main dry season food of camel and goat herds, is being overbrowsed and progressively eliminated. This decline is particularly evident in the 20 kilometres around permanent watering points (Kuchar, 1986).
There are a number of possibilities for the development and management of fodder resources both on farms, in rangelands and forest areas. Trials have been carried out to assess the potential for increased use of tree fodder as livestock feeds. Experiments with Leucaena fodder and cattle have shown that for beef fattening results are comparable with those of concentrated protein sources (when limited amounts are offered). Milk production was also improved although the Leucaena tainted the milk (Jones, 1979).
On rangelands, possibilities also exist for increasing livestock productivity through greater use of tree and shrub species. A number of species have been singled out with particular potential in this context, for example, Opuntia sp. and Atriplex nummalaria in arid areas of Africa (Kock, 1967), and Prosopis sp. in Latin America (Felker, 1979).
Improved management of rangelands may also include measures to control unpalatable woody species such as Calotropis procera, now common in many degraded Sahelian pastures, especially around boreholes, and Acacia reficiens, which has made large areas of Turkana, in Kenya, impenetrable to animals. In both cases, replacement by more palatable species would add considerably to the livestock carrying capacity of the area.
In introducing more trees to rangelands, one factor that has to be borne in mind is the trade-off between fodder production from woody species and that from the underlying grassy layer. A balance has to be struck between grasses, which give the greatest net productivity, and woody species, which are less productive but are better able to withstand drought. A combination that gives a high fodder yield during good years, may be disastrous if production drops dramatically during a dry year.
3.4.1
Trees and improvements to the soil
3.4.2
Nitrogen-fixing trees
3.4.3
Nutrient cycling in agroforestry systems
3.4.4
Possible detrimental effects of trees
Shifting cultivators and other farmers who depend on some kind of forest fallow have long recognized and depended upon the forest's (and tree's) ability to help improve soil conditions and thus indirectly crop yields. These effects are most pronounced in agroforestry systems in which trees or other woody perennials are grown in close association with agricultural crops. Such systems, exist in traditional forms in many parts of the world, as well as in a variety of new, experimental combinations (Nair, 1987a).
Over the last decade, a great deal of attention has been focused on the development potential of agroforestry systems (Sanchez, 1987). Agroforestry techniques can have a positive impact on crop production by improving the soil's physical properties, maintaining soil organic matter, and promoting nutrient cycling as well as reducing soil erosion and improving the micro-climate as was discussed in the previous chapter.
3.4.1 Trees and improvements to the soil
Under some circumstances, incorporating woody perennials on farmland can result in a marked improvement in soil fertility. There are several theories which explain the influence of trees on soil conditions, notably that the incorporation of trees may lead to:
* an increase in the organic matter content of the soil through addition of leaf litter, decaying roots and other plant parts;
* more efficient nutrient cycling within systems, and thus better utilization of nutrients that are either inherently present in the soil or externally applied;
* biological nitrogen fixation, and improved solubility of relatively unavailable nutrients, such as phosphate, as a result of the activity of micro-organisms in the tree root zone;
* an increase in the proportion of nutrients that are cycled through the plant layer, and therefore a decrease in nutrient loss through leaching;
* a moderating effect of additional soil organic matter on extremes in soil acidity and alkalinity, and, consequently, improved release and availability of nutrients such as phosphate and manganese that are sensitive to pH;
* increased activity of favourable micro-organisms in the root zone through improvement in the organic matter status and temperature of the soil;
* gradual improvement of the physical conditions of the soil - in permeability, water-holding capacity, aggregate stability, and soil temperature regimes.
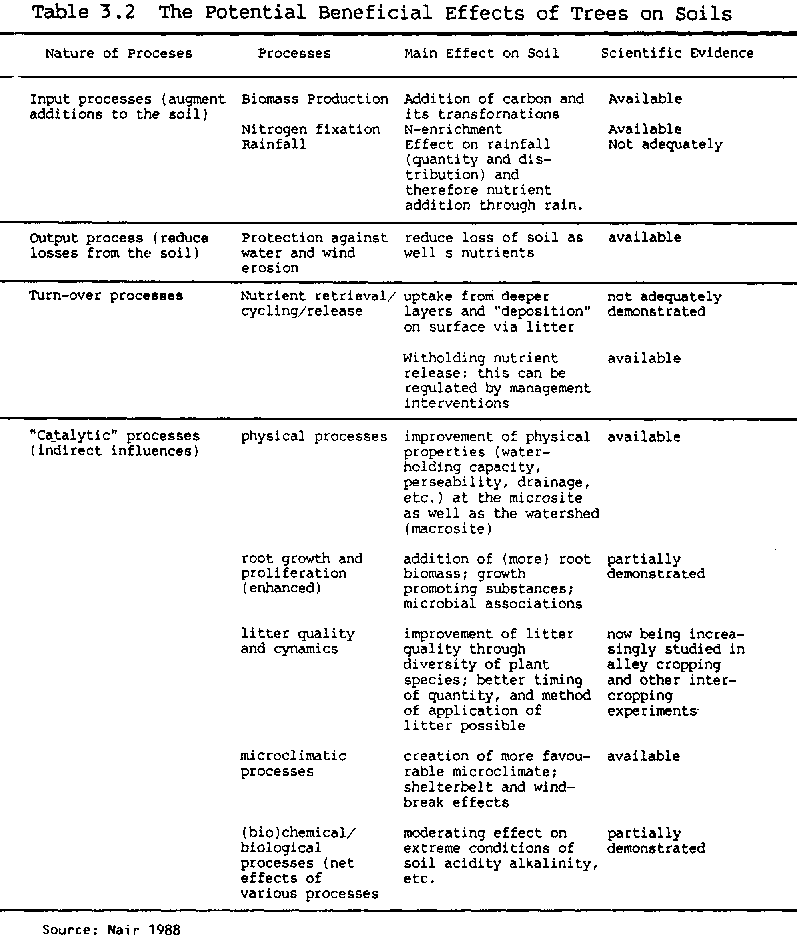
The relative significance of these different effects will vary greatly depending on the particular agroforestry system in question, and on local soil and site conditions. Many of these effects also take a considerable time to develop; trees cannot be expected to have a dramatic effect on soil fertility overnight. In addition, while these beneficial effects have been widely assumed, in practice not all of them have been scientifically demonstrated (Table 3.2 presents a summary of the current state of knowledge).
Table 3.2 the Potential Beneficial Effects of Trees on Soils
| Nature of Proceses | Processes | Main Effect on Soil | Scientific Evidence |
| Input processes (augment additions to the soil) | Biomass Production | Addition of carbon and its transformations . | Available |
| Nitrogen fixation Rainfall | N-enrichment Effect on rainfall (quantity and dis tribution) and therefore nutrient addition through rain | Available Not adequately | |
| Output process (reduce losses from the soil) | Protection against water and wind erosion | reduce loss of soil as well s nutrients | available |
| Turn-over processes | Nutrient retrieval/ cycling/release | uptake from deeper layers and "deposition" on surface via litter | not adequately demonstrated |
| Witholding nutrient release: this can be regulated by management interventions | available | ||
| "Catalytic" processes (indirect influences) | physical processes | improvement of physical properties (water- holding capacity, perseability, drainage, etc.) at the microsite as well as the watershed (macrosite) | available |
| root growth and proliferation (enhanced) | addition of (more) root biomass; growth promoting substances; microbial associations | partially demonstrated | |
| litter quality and cynamics | improvement of litter quality through diversity of plant species; better timing of quantity, and method of application of litter possible | now being increa singly studied in alley cropping and other inter cropping experiments | |
| microclimatic processes | creation of more favou- rable microclimate; shelterbelt and wind- break effects | available | |
| (bio)chemical/ biological processes (net effects of various processes | moderating effect on extreme conditions of soil acidity alkalinity, etc. | partially demonstrated |
One of most promising groups of tree species from the point of view of soil fertility are the nitrogen fixers. By virtue of their ability to capture atmospheric nitrogen and contribute nitrogen to the soil via leaf litter, or the release of root debris and nodules (root litter), this group of trees and shrubs can contribute significantly to the maintenance of soil fertility.
This ability is already made use of in many traditional agroforestry systems (Nair, 1987b; Dommergues, 1987). A number of points need to be borne in mind, however, when considering the potential for increased use of nitrogen-fixing trees:
* the ability of a particular species to fix nitrogen is highly site-specific and depends on climate, soil conditions and management practices;
* there is considerable variation in nitrogen-fixing ability between different provenances of the same species;
* effective nitrogen fixation requires the presence of the appropriate Rhizobium and Frankia strains in the root region;
* improvements in nitrogen fixation achieved in the laboratory, glasshouse or even in the nursery are not always easy to transfer to the field;
* even the least demanding nitrogen-fixing trees require other nutrients if they are to flourish, and these needs have to be met if their nitrogen-fixing potential is to be fully realised;
* the benefits of introducing nitrogen-fixing trees do not accrue instantaneously; the effects on soil fertility are often cumulative and may take several years to develop.
Thus, the fact that a particular tree is an effective nitrogen fixer under one set of conditions does not guarantee that it can be successfully transferred to another. To obtain the full benefits of nitrogen-fixing trees, a great deal of careful selection of species and provenances is often required, together with appropriate management to ensure that the necessary conditions for nitrogen fixation are provided. Nonetheless, nitrogen fixers can potentially contribute significantly to household food security in many farming situations.
3.4.3 Nutrient cycling in agroforestry systems
Agroforestry technologies (whether traditional or newly developed) improve soil conditions via more efficient cycling of nutrients. The potential soil benefits of these technologies depend greatly on local conditions and soil characteristics: tropical Alfisols and Andepts of moderate fertility appear to be particularly suited to agroforestry systems (Sanchez, 1987).
In one study in western Nigeria, for example, researchers found that planting Leucaena improved the regeneration of bush fallow on an Alfisol. After three years, during which Leucaena was cut annually and left as mulch, the Leucaena fallow resulted in significantly better soil conditions: higher effective cation exchange capacity and exchangeable calcium and potassium levels, compared to the bush fallow (duo and Lal, 1977).
The importance of site conditions in determining the effectiveness of agroforestry combinations has been clearly shown, however, in research carried out on alley cropping systems in different parts of the world. This system, which involves growing alternate rows of trees and crops, has proved highly successful in trials conducted by the International Institute for Tropical Agriculture (IITA), in Nigeria, using Leucaena leucocephala interplanted with maize and cowpeas. While alley cropping experiments in Nigeria over six years show a marked improvement in soil fertility (Kang et al, 1985), attempts to duplicate these experiments in a highly weathered, sandy Ultisol in the Amazon basin of Yurimaguas, Peru, were not as successful (TropSoils, 1986).
Although alley cropping works well in moderately fertile soils, current experience suggests that it will be necessary to use inputs such as lime, and possibly phosphorus, to allow successful establishment of alley cropping species and subsequent recycling of nutrients on infertile acid Ultisols and Oxisols (TropSoils, 1986). Further study is required before this particular agroforestry system can be considered widely applicable to the humid and sub-humid tropics. In addition, socio-economic aspects of the applicability of this particular system have not been well studied: i.e. seasonal labour requirements and availability, availability and access to inputs, access to land for tree planting and other tenure issues, and management requirements. In many regions the right socio-economic conditions may not exist for the development of alley-cropping.
3.4.4 Possible detrimental effects of trees
The discussion thus far has highlighted only the benefits that trees provide. The effects of trees on crops are not always positive. If the wrong species are chosen, or if they are planted in an inappropriate manner - for example, at too close a spacing - then trees can have a variety of adverse effects on crops grown in their immediate vicinity. These effects include:
* fast growing trees place a heavy demand on soil moisture; where crop growth is being limited by moisture availability, competition with trees will reduce crop yields;
* uptake of nutrients by trees may deprive adjacent crops of nutrients (although litter fall, and root biomass may compensate for this to some extent in the long run);
* certain trees have adverse chemical and biological effects on nearby plants as a result of acidification, alleopathy, production of toxic exudates, or by providing a habitat for crop nests:
* shading and changes in spectral quality of light can have a detrimental effect on the growth of crop species in close proximity to trees.
Once again, these effects are highly site-specific and depend not just on the combination of species being used but also on the way in which they are arranged, and the management methods used. Successful agroforestry systems are those that maximise positive interactions, while minimising the negative ones. In traditional agroforestry systems the most effective combinations have often been worked out over generations. With the new techniques currently being developed, it is only by a great deal of careful research and on-farm trials that the management and species optima can be approached. In addition, to the physical problems associated with incorporating trees into farming, there are many socio-economic factors which influence the viability of agroforestry technologies for a particular household or community. These are discussed in the following chapter.
3.5 Food production from mangroves
3.5.1
Mangroves: support for coastal fisheries
3.5.2
Additional food products from mangroves
3.5.3
Pressures on mangrove ecosystems
Mangrove forests are unique ecosystems, considered separately here because they contribute to food security in special ways, most notably supporting coastal fisheries. Mangrove forests exist along coastlines in a number of tropical and subtropical areas. Their unique flora is specially adapted to periodic submersion by salt water. They produce a range of plant foods and provide a habitat and breeding ground for a large number of marine animals. In addition they provide a buffer to coastal communities and their cultivated lands against sea storms (as was discussed in the previous chapter).
The total area of mangroves worldwide is estimated at between 160,000 and 170,000 square kilometres (Saenger, 1983). The largest areas are in Brazil, followed by Indonesia, Australia, Nigeria and Malaysia (Hamilton and Snedaker, 1984). They exist in a dynamic state, with their area gradually being extended through the process of siltation, but also being subjected to periodic destruction caused by erosion and violent storms.
3.5.1 Mangroves: support for coastal fisheries
Mangrove forests have a major role in supporting offshore fisheries, and thus in protecting a major food source for many coastal populations. In the Pichavaram mangrove in southern India, for example, 74% of the penaeid prawns caught in adjacent coastal waters use the mangrove as nursery grounds (Krishnamurthy 1984). In the Gulf of Mexico, it is estimated that 90% of the commercial catch and 70% of the recreational catch are dependent on mangrove estuaries for some part of their life cycle, either during the breeding stage or as larvae, juveniles or adults. Most of the information on mangrove-dependent fish and animals focuses on important commercial species including: bream, mullet, milkfish, mojarras, snooks, barramundi, seatrout, snapper, drum, croaker, grouper and tarpon (Hamilton and Snedaker, 1984). There are undoubtedly many little known species which also depend on these areas and provide a staple supply of food for nearby communities.
Large quantities of fish, shrimp, oysters, crabs, cockles and other marine animals are caught in mangroves themselves. The total annual catch including fish, molluscs, crabs and shrimp is estimated at around one million tons, slightly over 1% of the total world fish catch (Kapetsky, 1987). Besides the contribution they make to local diets, mangrove fisheries provide employment for up to half a million people. In most mangrove areas, the income generated from fishery products is several times greater than that from forestry.
Oysters, snails, mussels and other molluscs are also cultured in some mangrove areas. The techniques used range from inexpensive ''spat collectors", where oysters are harvested from aerial branches, to special rafts as used in the Philippines.
In recent years there has been an increasing shift towards the use of ponds for rearing fish and shrimp - commonly referred to as aquaculture. These range from simple ponds which rely on the movement of the tides to bring in seawater and fresh nutrients, to more elaborate aquaculture systems involving separate hatcheries and nurseries, provision of feeds, and use of pumping to regulate water flows. Even these systems, however, rely to a certain extent on mangrove areas for nutrients as well as fry recruitments (Christensen, 1983).
3.5.2 Additional food products from mangroves
A wide range of other food products are also obtained from mangroves, either cultivated or collected from the wild.
* Honey is collected from many mangrove forests. Total production from wild hives in the Sundarbans in Bangladesh forest, for example, was estimated at 263,000 kg in 1983/4 (Masson, 1984). In Cuba, up to 30,000 hives are moved each year, following the flowering season of Avicennia, which develops in April in the Southwest and lasts until August in the North and East of the island.
* Algae are increasingly cultivated in some countries. In Thailand, Gracilaria is grown, the best sites being shorelines with sandy bottoms. In the Philippines, algae is cultivated as a high value crop for export to Japan (Deveau and Castle, 1976).
* Fruits such as from the Nipa palm are collected from some mangrove forests and provide an important contribution to local diets.
* Salt is produced by evaporating seawater in many mangrove areas. In Pakistan, for example, on the Gulf of Kutch, there are 15,000 hectares of saltflats at the back of or in depleted low scrub mangroves. Thailand produces over 400,000 tons of salt a year in saltflats converted from mangroves. Some saltflats are turned into fishponds in the wet season (Hamilton and Snedaker, 1984).
* Leaves, especially from Rhizophora, are gathered from some mangrove forests and provide a high protein animal feed. In Iran, the United Arab Emirates and Pakistan, camels are traditionally allowed to graze in mangrove areas (Kulkarni and Junagad, 1959).
* Emergency foods can be obtained from some mangrove plants. Avicennia can be eaten if boiled several times, and Pacific islanders use Bruquiera gymnorrhiza hypocotlys baked into a sort of bread, after peeling to remove excess tannin. A wide variety of traditional medicines are also derived from mangrove plants.
3.5.3 Pressures on mangrove ecosystems
Although relatively undisturbed mangroves still exist in some countries, in recent years mangroves have been increasingly affected by human pressures. Large areas have been converted for other uses, and many of the areas that remain are being gradually degraded.
Land reclamation for agriculture is one of the main reasons for the loss of mangrove areas, although high salinity and the tendency for soils to become acidified make it a lengthy and often problematic operation. Urban expansion is another factor; many major coastal cities are sited partly or entirely on mangrove grounds - Miami, Panama City, Guayaquil, Sao Luis, Cotonou, Bombay, Jakarta, and Manila are just some examples. As these cities have grown, increasing areas of mangroves have been reclaimed. In addition, conversion of mangroves to aquaculture ponds has had a major effect in some countries.
Wholesale destruction, or the conversion to other uses, pose an obvious threat to mangrove areas. What is less apparent is the gradual degradation of mangroves through various human influences. In the long run, however, these may be even more damaging.
These problems relate to a general lack of land use planning for mangrove areas. Construction of fish and shrimp ponds often result in excessive destruction of mangrove areas and the degradation of water and nutrient quality. The uncontrolled harvesting of firewood and poles, and the lack of replanting pose another problem. In addition, the various forms of pollution urban wastes, insecticides, wastes from sugar cane and other food processing industries, heavy metals from mining, oil spills, thermal pollution from power stations - all induce degradation of mangroves. Finally, the construction of large dams can also have a significant impact on mangrove survival as they affect river flows and sediment loads.
In practice, pinpointing the causes of mangrove degradation can be extremely difficult as all of these factors are interconnected. While destruction and degradation of mangroves can be expected to have a variety of deleterious effects, there are no simple relationships linking, for example, loss of mangrove areas with reduced offshore fish catches.
There is, however, a clear need for more rational and sustainable management of the remaining mangrove areas - to preserve the important role they play both in providing food, and in supplying other locally important products and benefits.
{kind=link}